



Pratique | Débuter
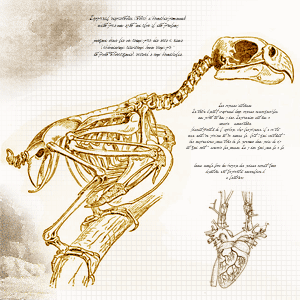
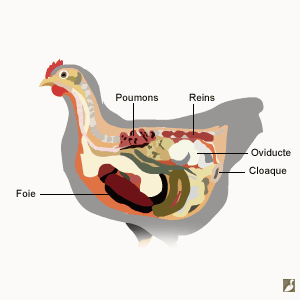
Le système digestif des oiseaux
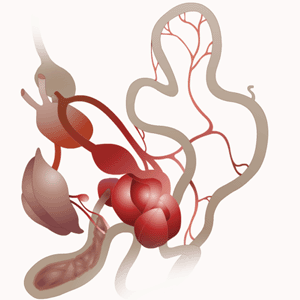
Schéma général du système digestif d’un oiseau.
Schéma : Ornithomedia
Introduction
Le système digestif des oiseaux a pour but de convertir la nourriture en matières premières indispensables au fonctionnement de l’organisme et à la création des cellules. Il prend en charge la nourriture, la décompose en molécules nutritives, les fait passer dans le flux sanguin et débarrasse le corps des substances non digestes.
Dans cet article, nous décrivons le système digestif des oiseaux et leur mode de digestion.
Abstract
The body is divided into different systems and their organs. The digestive system of birds, as for others animals, converts food into raw materials that build and fuel the body’s cells. It takes in food, breaks down nutrient molecules, absorbs them into the blood stream and rids the body of the indigestible remains.
In this article, we present you the different organs, from the Esophagus and Crop to the Cloaca, their functions, and we present the mecanism of the digestion.
Poursuivez la lecture de cet article, en vous abonnant dès maintenant !
Découvrez les Archives d’Ornithomedia.com
Pour seulement 10,00 €TTC/an (ou 6,00 € les 6 mois)
Profitez de plusieurs centaines d’articles en accès illimité et sans aucun engagement.
Compléments
Ouvrages recommandés
- The Inner Bird: Anatomy and Evolution (2008) de Gary W. Kaiser
- Biologie animale – Les Cordés – 9ème édition – Anatomie comparée des vertébrés: Anatomie comparée des vertébrés (2009) d’André Beaumont (Auteur), Pierre Cassier (Auteur), Daniel Richard





Aucun commentaire sur ce sujet
Participer à la discussion !