



Magazine | Interviews
Tammy Steeves et la « redécouverte » du Fou de Tasmanie
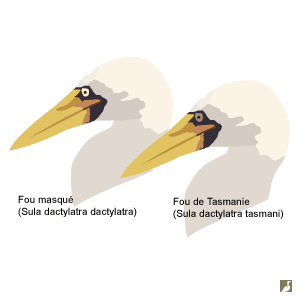
Fous masqué (Sula dactylatra dactylatra) et de Tasmanie (S. d. tasmani) : ce dernier a un iris sépia et a des ailes légèrement plus longues.Schéma : Ornithomedia.com
Introduction
On croyait que le Fou de Tasmanie avait été exterminé sur l’île de Norfolk par les Polynésiens vers 1200 puis sur l’île de Lord Howe au 18ème siècle par des marins européens affamés; mais les chercheurs Tammy Steeves, Marie Hale et Richard Holdaway de l’Université de Canterbury (Nouvelle-Zélande) ont démontré qu’il vivait toujours et correspondait en fait à la sous-espèce actuelle fullagari du Fou masqué (Sula dactylatra) ! Le Fou de Tasmanie porte désormais un nouveau nom scientifique, Sula dactylatra tasmani. Les résultats de cette étude ont été publiés le 12 août 2009 dans le journal Biology Letters.
Cette sous-espèce niche sur trois îles dispersées dans la Mer de Tasmanie: Norfolk, Lord Howe et Kermadec. Sula dactylatra tasmani a un iris de couleur sépia et non pas jaune comme la sous-espèce nominale, et ses ailes sont plus longues de 10 % environ.
Tammy Steeves, professeur à la School of Biological Sciences, (Université de Canterbury, Nouvelle-Zélande), a répondu à nos questions.
Abstract
Three University of Canterbury academics have helped rediscover a seabird thought to have been driven to extinction by hungry European sailors in the late 18th century. Dr Tammy Steeves, Dr Marie Hale and Adjunct Professor Richard Holdaway (Biological Sciences) are part of a team of scientists from across New Zealand and Australia who have used an innovative multidisciplinary approach to resolve the taxonomic status of the “extinct” Tasman booby (Sula tasmani). Using classical palaeontological data combined with ancient and modern DNA data, the biologists discovered that the Tasman booby (Sula tasmani) turns out be a subspecies of a living species, the masked booby (Sula dactylatra fullagari). And now these charismatic seabirds have a now new scientific name, Sula dactylatra tasmani.
Poursuivez la lecture de cet article, en vous abonnant dès maintenant !
Découvrez les Archives d’Ornithomedia.com
Pour seulement 10,00 €TTC/an (ou 6,00 € les 6 mois)
Profitez de plusieurs centaines d’articles en accès illimité et sans aucun engagement.
Compléments
À lire aussi sur Ornithomedia.com
- Croisière ornithologique mouvementée sur la mer de Tasman
- À la découverte des îles Snares, Auckland et Campbell
Ouvrages recommandés
- Birds of Australia de Ken Simpson (Auteur) et Nicolas Day (Auteur)
- Field Guide to New Zealand Seabirds de Brian Parkinson
- Albatrosses de Terence Lindsay
- Albatrosses, Petrels and Shearwaters of the World de Derek Onley (Auteur) et Paul Scofield (Auteur)
Source
Tammy E. Steeves, Richard N. Holdaway, Marie L. Hale, Emma McLay, Ian A. W. McAllan, Margaret Christian, Mark E. Hauber, et Michael Bunce (2009). Merging ancient and modern DNA: extinct seabird taxon rediscovered in the North Tasman Sea. Biology Letters. Date de mise en ligne: 12 août 2009. http://rsbl.royalsocietypublishing.org/content/early/2009/08/07/rsbl.2009.0478
Aucun commentaire sur ce sujet
Participer à la discussion !